Про воду и расщепление жиров (липидов). Сложновато, но понятно! Кафедра биохимии
Расщепление липидов в желудочно-кишечном тракте.
Липиды, поступающие с пищей, крайне гетерогенны по своему происхождению. В желудочно кишечном тракте они в значительной мере расщепляются до составляющих их мономеров: высших жирных кислот, глицерола, аминоспиртов и др. Эти продукты расщепления всасываются в кишечную стенку и из них в клетках кишечного эпители синтезируются липиды, свойственные человеку. Эти видоспецифические липиды далее поступают в лимфатическую и кровеносную системы и разносятся к различным тканеям и органам. Липиды, поступающие из кишечника во внутреннюю среду организма обычно называют экзогенными липидами.
Процесс расщепления пищевых жиров идет в основном в тонком кишечнике. В пилорическом отделе желудка, правда, выделяется липаза, но рН желудочного сока на высоте пищеварения составляет 1,0 - 2,5 и при этих значениях рН фермент малоактивен. Принято считать, что образующиеся в пилорическом отделе желудка жирные кислоты и моноглицериды далее участвуют в эмульгировании жиров в двенадцатиперстной кишке. В желудке под действием протеиназ желудочного сока происходит частичное расщепление белковых компонентов липопротеидов, что в дальнейшем облегчает расщепление их липидных составляющих в тонком кишечнике.
Поступающие в тонкий кишечник липиды подвергаются действию ряда ферментов. Пищевые триацилглицерины (жиры) подвергаются действию фермента липазы, поступающей в кишечник из поджелудочной железы. Эта липаза наиболее активно гидролизует сложноэфирные связи в первом и третьем положении молекулы триацилглицерина, менее эффективно она гидролизует сложноэфирные связи между ацилом и вторым атомом углерода глицерола. Для проявления максимальной активности липазы требуется полипептид - колипаза, поступающий в двенадцатиперстную кишку, по-видимому, с соком поджелудочной железы. В расщеплении жиров участвует также липаза, выделяемая стенками кишечника, однако, во-первых, эта липаза малоактивна; во-вторых, она преимущественно катализирует гидролиз сложноэфирной связи между ацилом и вторым атомом углерода глицерола.
При расщеплении жиров под действием липаз панкреатического сока и кишечного сока образуются преимущественно свободные высшие жирные кислоты, моноацилглицерины и глицерол. В то же время, образующаяся смесь продуктов расщепления содержит и некоторое количество диацилглицеринов и триацилглицеринов. Принято считать, что лишь 40-50% пищевых жиров расщепляется полностью, а от 3% до 10% пищевых жиров могут всасываться в неизмененном виде.
Расщепление фосфолипидов идет гидролитическим путем при участии ферментов фосфолипаз, поступающих в двенадцатиперстную кишку с соком поджелудочной железы. Фосфолипаза А1 катализирует расщепление сложноэфирной связи между ацилом и первым атомом углерода глицерола. Фосфолипаза А2 катализирует гидролиз сложноэфирной связи между ацилом и вторым атомом углерода глицерола. Фосфолипаза С катализирует гидролитический разрыв связи между третьим атомом углерода глицерола и остатком фосфорной кислоты, а фосфолипаза Д ДД сложноэфирной связи между остатком фосфорной кислоты и остатком аминоспирта.
В результате действия этих четырех ферментов фосфолипиды расщепляются до свободных жирных кислот, глицерола, фосфорной кислоты и аминоспирта или его аналога, например, аминокислоты серина, однако часть фосфолипидов расщепляется при участии фосфолипазы А2 только до лизофосфолипидов и в таком виде может поступать в стенку кишечника.
Сложные эфиры холестерола расщепляются в тонком кишечнике гидролитическим путем при участии фермента холестеролэстеразы до жирной кислоты и свободного холестерола. Холестеролэстераза содержится в кишечном соке и соке поджелудочной железы.
Все ферменты, принимающие участие в гидролизе пищевых липид растворены в водной фазе содержимого тонкого кишечника и могут действовать на молекулы липидов лишь на границе раздела липид/вода. Отсюда, для эффективного переваривания липидов необходимо увеличение этой поверхности с тем, чтобы большее количество молекул ферментов участвовало в катализе. Увеличение площади поверхности раздела достигается за счет эмульгирования пищевых липидов ДД разделения крупных липидных капель пищевого комка на мелкие. Для эмульгирования необходимы поверхностно-активные вещества - ПАВы, представляющие собой амфифильные соединения, одна часть молекулы которых гидрофобна и способна взаимодействовать с гидрофобными молекулами поверхности липидных капель, а вторая часть молекулы ПАВов должна быть гидрофильной, способной взаимодействовать с водой. При взаимодействии липидных капель с ПАВами снижается величина поверхностного натяжения на границе раздела липид/вода и крупные липидные капли распадаются на более мелкие с образованием эмульсии. В качестве ПАВов в тонком кишечнике выступают соли жирных кислот и продукты неполного гидролиза триацилглицеринов или фосфолипидов, однако основную роль в этом процессе играют желчные кислоты.
Желчные кислоты, как уже упоминалось, относятся к соединениям стероидной природы. Они синтезируются в печени из холестерола и поступают в кишечник вместе с желчью. Различают первичные и вторичные желчные кислоты. Первичными являются те желчные кислоты, которые непосредственно синтезируются в гепатоцитах из холестерола: это холевая кислота и хенодезоксихолевая кислота. Вторичные желчные кислоты образуются в кишечнике из первичных под действием микрофлоры: это литохолевая и дезоксихолевая кислоты. Все желчные кислоты поступают в кишечник с желчью в коньюгированных формах, т.е. в виде производных, образующихся при взаимодействии желчных кислот с гликоколом или таурином.
Кроме наличия ПАВов для эмульгирования имеют значение постоянное перемешивание содержимого кишечника при перистальтике и образование пузырьков СО2 при нейтрализации кислого содержимого желудка, поступающего в двенадцатиперстную кишку, бикарбонатами сока поджелудочной железы, поступающего в этот же отдел тонкого кишечника.
Всасывание продуктов переваривания липидов
В стенку кишечника легко всасываются вещества, хорошо растворимые в воде. Из продуктов расщепления липидов к ним относятся, например, глицерол, аминоспирты и жирные кислоты с короткими углводородными радикалами (до 8 - 10 атомов "С"), натриевые или калиевые соли фосфорной кислоты. Эти соединения из клеток кишечника обычно поступают непосредственно в кровь и вместе с током крови транспортируются в печень.
В то же время большинство продуктов переваривания липидов: высшие жирные кислоты, моно- и диацилглицерины, холестерол, лизофосфолипиды и др. плохо растворимы в воде и для их всасывания в стенку кишечника требуется специальный механизм. Перечисленные соединения, наряду с желчными кислотами и фосфолипидами, образуют мицеллы. Каждая мицелла состоит из гидрофобного ядра и внешнего мономолекулярного слоя амфифильных соединений, расположенных таким образом, что гидрофильные части их молекул контактируют с водой, а гидрофобные участки ориентированы внутрь мицеллы, где они контактируют с гидрофобным ядром. В состав мономолекулярной амфифильной оболочки мицеллы входят преимущественно фосфолипиды и желчные кислоты, сюда же могут быть включены молекулы холестерола. Гидрофобное ядро мицеллы состоит преимущественно из высших жирных кислот, продуктов неполного расщепления жиров, эфиров холестерола, жирорастворимых витаминов и др.
Благодаря растворимости мицелл возможен транспорт продуктов расщепления липидов через жидкую среду просвета кишечника к щеточной каемке клеток слизистой оболочки, где эти продукты всасываются. В норме всасывается до 98% пищевых липидов.
Поступившие в энтероциты мицеллы разрушаются. Всосавшиеся продукты расщепления экзогенных липидов превращаются в энтероцитах в липиды, характерные для организма человека, и далее они поступают во внутреннюю среду организма. Высвободившиеся при распаде мицелл желчные кислоты из энтероцитов или поступают обратно в кишечник, или же поступают в кровь и через воротную вену оказываются в печени. Здесь они улавливаются гепатоцитами и вновь направляются в желчь для их повторного использования.
Такая энетро-гепатическая циркуляция желчных кислот, обеспечивающая их неоднократное использование, позволяет существенно снизить объем их ежесуточного синтеза. Общий пул желчных кислот в организме составляет 2,8 - 3,5 г. Они совершают 5-6 оборотов в сутки. Конечно, часть желчных кислот ежесуточно теряется с калом. Эти потери составляют по разным оценкам от 0,5г до 1,0 г в сутки. Потери восполняются их синтезом из холестерола.
Кстати, при нарушении поступления желчных кислот в кишечник в результате закупорки желчевыводящих путей больше страдает процесс всасывания продуктов расщепления липидов в стенку кишечника, нежели механизм переваривание липидов. Именно поэтому каловые массы у таких больных содержат большое количество солей высших жирных кислот, а не неизмененных липидов. Естественно, что в этой ситуации нарушается и всасывание жирорастворимых витаминов, так как они поступают в энтероциты также в составе мицелл.
1. Липиды - природные органические соединения (крайне гетерогенны по своей химической структуре) общими свойствами которых является: низкая растворимость в воде и хорошая растворимость в аполярных растворителях таких как хлороформ, жидкие углеводороды и др. Липидная фракция клеток - все то, что можно извлечь из клеток с помощью полярных растворителей.К липидам относятся соединения имеющие весьма различную химическую природность: например холистерол - циклически непридельный одноатомный спирт. Сравните его со структурой триацилглицерина Сходство между ними практически нет. Это крайне различные соединения.Согласно современной классификации все липиды делятся на 4 больших группы:
Жирные кислоты - это алифатические карбоновые кислоты число атомов в которых может достигать 22-24. Основная масса жирных кислот входящих в организм человека и животных имеет четное число атомов углерода, что связано с особенностями их синтеза. Дело в том, что синтез идет путем тотарного удлинения углеродной цепочки.
Жирные кислоты как правило имеют неразветвленную углеводородную цепь
Соединения смешанной природы.К этой группе относятся соединения сложной химической природы одним из компонентом которых является липид. К таким соединениям относится например а) Липополисахарида б) Липоаминокислоты в) Липопротеиды (сегодня считают их надмолекулярными комплексами) Они принимают участие в транспорте липидов кровью. уЪтиды нерастворимы в воде и не могут транспортироваться в водной фазе. Даже клеточные мембраны представляют собой липопротеидные надмолекулярные структуры.
2. Переваривание липидов. Всасываний продуктов переваривания липидов.В стенку кишечника легко всасываются вещества хорошо растворимые в воде. Из продуктов расщепления липидов к ним относятся такие как глицерол, аминоспирты, жирные кислоты с количеством углеродных атомов не более 10, а так же натриевые соли фосфорной кислоты. Эти соединения из клеток.кишечника обычно лоступают непосредственно в:кровь и с током.крови транспортируются в лечень. В то же время большинство продуктов переваривания липидов - это высшие жирные кислоты, моно- и диацилдглицерины, лизофосфолипиды, холистерол и др, плохо растворимы в воде и для всасывания их в стенку кишечника требуется специальный механизм.
Перечисленные соединения наряду с желчными кислотами и фосфолипидами образуют мицелу. Мицела состоит из гидрофобного ядра и внешнего мономолекулярного слоя амфифильных соединений. Эти амфифильные соединения расположены таким образом, что гидрофильные их части контактируют с водой, т.е. направлены кнаружи, а гидрофобные участки ориентированы во внутрь мицелы, где контактируют с гидрофобным ядром. В состав наружной оболочки мицелы входят преимущественно фосфолипиды и желчные кислоты сюда же может входит и холистерол, поскольку это сепирт. Гидрофобное ядро мицелы состоит из высших жирных кислот, продуктов неполного расщепления жиров, эфиров холестерина, жирорастворимых витаминов.
Благодаря растворимости мицел эти продукты всасываются путем эндоцитоза. В норме у нас всасывается до 98% пищевых липидов, 2% выбрасываются. Ресинтез липидов в кишечной стенке. В кишечной стенки всосавшиеся ацилглицерины распадаются под действием тканевых липаз с образованием свободных жирных кислот и глицерола. Часть моноацилглицеринов может превращаться в триацилглицерины без предварительного расщепления. Это так называемый моноацилглицериновый путь ресинтеза. Все высшие жирные кислоты всосавшиеся из кишечника используются в энтероцитах для ресинтеза различных липидов. Но перед тем как включаться в различные липиды, высшие жирные кислоты должны быть активированы.
Процесс активации высших жирных кислот состоит из 2 этапов: " I этап. За счет взаимодействия жирных кислот и использования специального фермента образуется ациладенилаты. Кроме того образуется второй продукт - пирофосфат, который немедленно расщепляется до фосфорной кислоты (так называемый термодинамический контроль направления процесса)
2 этап. Происходит образование активной жирной кислоты соединенной с КоА и высвобождение АМФ. Образование ацилКоА катализируется специальным ферментом, причем он катализирует и первую и вторую реакцию ацилКоАсинтетаза (тиокиназа).
В ходе активации высшей жирной кислоты АТФ распадается до АМФ и 2 остатков фосфорной кислоты, таким образом активация обходится клетке в 2 макроэргических эквивалента. Во всех своих превращениях в клетках жирные кислоты участвуют в активированной форме.
3. Мобилизация триглицеридов жировой ткани и проблемы транспорта высших жирных кислот. Постадсорбционном периоде (когда между приемами пищи длительный интервал) идет мобилизация энергетических ресурсов организма в том числе мобилизация триглицеридов жировой ткани. Образующиеся в ходе мобилизации высшие жирные кислоты через мембрану липоцитов поступают в кровяное русло и в комплексе с альбуминами переносятся током крови в различные органы и ткани. Там жирные кислоты проникают через наружную клеточную мембрану внутрь клеток и связываются со специальным так называемым Z-белком. В комплексе с этим внутриклеточным переносчиком жирные кислоты перемещаются в цитозоле к месту их использования. Концентрация неэтерифицированных или иначе свободных жирных кислот в плазме крови натощак составляет величину 0,56-0,58 млмоль/л. Жирные кислоты очень быстро обмениваются в крови, время их полужизни в русле крови составляет около 4 мин. За сутки с током крови переноситься примерно 150 гр свободных жирных кислот. Эта величина превышает величину суточного поступления липидов в организме. Это свидетельствует о том, что значительная часть транспортируемых кровью высших жирных кислот является продуктом их биосинтеза из углеводов или углеродного скелета аминокислот. В условиях длительно интенсивной работы требующей больших энергозатрат жирные кислоты, поступающие из жировых депо становятся основным видом энергетического топлива. Значение их как энергетического топлива еще более возрастает при недостатке глюкозы в органах и тканях, что характерно для сахарного диабета или голодания. Однако на пути эффективного использования высших жирных кислот клетками встает так называемый диффузионный барьер.Что это за барьер?Суть этого явления заключается в том, что высшие жирные кислоты на своем пути из кровяного русла в клетки должны пройти через гидрофильную фазу межклеточной среды. Но они нерастворимы в воде и поэтому скорость движения через межклеточную среду крайне ограничена. Выходом из положения является преобразование жирных кислот в печени в соединения с небольшой молекулярной массой которые растворимы в воде.
Это-такиее|соединения как ацегоуксусные и B-гидроксимасляные кислоты. Эти соединения из печени „щять-же поступают в кровь, а затем идут в клетки тканей, но для этих молекул диффузионного барьера не существует, поэтому они служат эффективным энергетическим топливом. Эти соединения получили название - ацетоновые тела. К ацетоновым телам относится и сам ацетон (диметилкетон). В то же время в гепатоциты высшие жирные кислоты поступают минуя диффузионный барьер потому, что гпатоциты в печеночных синусах непосредственно контактируют с кровью.
4. Процессы окислительного расщепления липидов в тканях. В-окисление насыщ жирных кислот, особенности окисления ненасыщенных жирных кислот и с нечетным количеством атомов углерода, окисление глицерина, синтез и окисление ацетоновых тел. Окисление глицерина- Глицерина в триглицеридах содержится 3-5%. Свободный глицерол, образующийся при гидролизе резервных триглицеридов или поступивший из кишечника человека, в клетках может окисляться.__
При полном окислении глицерина в клетке синтезируется 23 молекулы АТФ. 1 молекула АТФ израсходуется на активацию глицерола: на его фосфорилирование, таким образом полный выход АТФ при окислении глицерола составит 22 молекулы АТФ.На долю глицерина в триглицеридах приходиться 3-5% поэтому существенного вклада в обеспечении энергией клеток различных тканей и органов окисление глицерина вносить не может. синтез и окисление ацетоновых тел.- Жирные кислоты поступающие в гепатоциты, активируются и подвергаются в-окислению с образованием ацетилКоА. Именно этот ацетилКоА используется для синтеза ацетоновых тел, согласно схеме.__
Последняя реакция - лиазная (катализирует фермент ГМГ-лиаза), происходит отщепление ацетилКоА и образование 4-х углеродной молекулы - ацетоацетата.__
Из ацетоуксусной кислоты спонтанно, чаще всего, или иногда за счет декарбоксилазы происходит отщепление карбоксильной группы в виде углекислого газа и образуется ацетон.
Образовавшиеся ацетоновые тела поступают из гепатоцитов в кровь и разносятся к клеткам. Процесс синтеза ацетоновых тел идет постоянно и ацетоновые тела всегда присутствуют в крови в концентрации ЗОмг/л. При голодании их содержание может увеличиваться до 400-500 мг/л. Еще больше концентрация при сахарном диабете в тяжелой форме до 3000-4000 мг/л.
Ацетоновые тела в норме хорошо утилизируются клетками периферических тканей, в особенности это касается скелетных мышц и миокарда. Скелетные мышцы и миокард значительную часть нужной им энергии получают за счет окисления ацетоновых тел.
5. Представления о биосинтезе выш жирных кислот, фосфолипидов, холестерине. В органах и тканях человека синтезируются почти все необходимые жирные кислоты за исключением полиеновых (вжк). Исходным соединением для синтеза (вжк) является ацетилКоА, который в клетках может образовываться из различных соединений. Преимущественно используется ацетилКоА,образующийся при окислении моносахаридов.Одновременно используется ацетилКоА, который образуется при распаде углеродных скелетов аминокислот. Синтез (вжк) может протекать в клетках, однако основная масса соединений этого класса синтезируется в печени и жировой ткани. Важнейшим субстратом, продукты метаболизма которого используются для синтеза липидов, является глюкоза АцетилКоА используемый при липогенезе образуется в основном в матрикее митохондрий, Синтез высших жирных кислот идет в цитозоле Внутренняя мембрана митохондрий непроницаема для ацетилКоА поэтому существует система (челночная) транспорта ацетильных остатков из матрикса митозхондрий в цитозоль.
Синтез фосфолипидов Все необходимые организму фосфолипиды могут синтезироваться в его клетках, причем в клетках могут функционировать несколько различных метаболических путей биосинтеза глицерофосфолипидов. Синтез лицитина А) при реакции ацилирования образуется фосфотидная кислота далее за счет фермента фосфотазы фосфотиднойкислоты образуется диглнцерид Б) Параллельно идет активация аминоспиртов (при наличии свободных аминоспиртов в клетке) далее идетактивация холина с образованием его производного - ЦДФхоли (активированный) он может включаться всинтезх.в) Диглицерид+ЦДФхолин - трансферазная реакция обеспечивает образование фосфотидилхолина и отщепляется ЦМФ, который затем за счет энергии АТФ может превращаться в ЦДФ. Алътернатнвным вариантоу синтеза может быть синтез с промежуточным образованием фосфотидной кислоты, но уже активированной. Сама фосфотидная кислота образуется при помощи ацилирования фосфоглицерина. Далее она взаимодействует с цитидин-3-фосфатом с образованием активной фосфотиднои кислоты (ЦДФ-фосфотидная кислота). Далее идет превращение в фосфотидилсерин или в инозитолфосфатид В том и другом случае происходит отщепление ЦМФ
Синтез Холистерола Он синтезируется в клетках из двух углеродных группировок ацетилКоА. Процесс синтеза включает в себя 35 последовательных реакций и может быть разбит на 4-5 этапов:1 этап - образование из ацетилКоА мевалоновой кислоты. 2 этап - -образование из мевалоновой кислоты активированных 5 углеродных группировок, изопреноидные группировки (это изопентилпирофосфат, димитнлаллилпирофосфат - активные изопреноидные группировки). 3 этап - конденсация изопреноидных группировок с образованием сквалена.4 этап - циклизация сквалена в ланостерин.5 этап - преобразование ланостерина в холистерол.
6. Биосинтез триглицеридов.Триглицериды синтезируются а клетках практически всех органах и тканей в качестве резервных питательных веществ, однако необходимо подчеркнуть, что синтез интенсивный с наибольшей интенсивностью идет в клетках печени и клетках жировой ткани
Что необходимо для синтеза триглицеридов? Для синтеза необходимы ВЖК и глицерол ВЖК поступают в клетки или из плазмы крови или же синтезируются на месте, т е. в клетках непосредственно, из ацетилКоА.
Глицерол может поступать так же из плазмы крови, однако основным источником глицерина для синтеза триглицеридов и даже фосфолипидов в клетках служит промежуточный продукт распада углеводов -фосфодиоксиацетон
ВЖК участвуют в синтезе триглицеридов в виде своих активированных производных - ацилКоА. Необходимый для синтеза фосфоглицерин образуется или путем восстановления фосфодиоксиацетона, или же за счет фосфорилирования свободного глицерола Эта реакция катализируется АТФ зависимой глицеролкиназой
После образования фосфоглицерина за счет двух последовательных реакции ацилирования образуется фосфотидная кислота, затем от нее отщепляется остаток фосфорной кислоты и ооразуется диглицерид
И наконец в ходе последней реакции ацилирования образуется триглицерид.__
В постадсорбцеонном периоде происходит мобилизация резервных триглицеридов. Расщепление триглицеридов в клетках идет под действием ферментов липаз. По крайней мере в липоцитах жировой ткани при распаде триглицеридов работают три различных фермента.Вначале при участии триацилглицероллипазы происходит гидролиз одной сложной эфирной связи образуется диглицерид и высвобождается свободная жирная кислота. Затем под действием второго фермента так же происходит гидролиз второй сложной эфирной связи и высвобождается еще одна ВЖК остается моноглицерид. С участием последней липазы происходит расщепление моноглнцерида на глицерол и ВЖК, фермент -моноацилглицероллипаза.
7. Липид транспортная система плазмы крови. Липиды практически нерастворимы в воде, в связи с этим возникает проблема транспорта в организме человека Основные показатели содержания липидов в плазме крови натощак для здорового человека в состоянии покоя Содержание общих липидов в крови составляет от 3 до 8,5 гр/л Содержание свободных жирных кислот в плазме крови 0,68-0,88 млмоль/л Общий холистерол 3,5 - 6,8 млчоль/л^триглицериды 0,6 - 2,3 млмоль/л, общие фолсфолипиды 2 - 4,7 млмлоль/л Все липиды присутствующие в крови входят в состав смешанных надмолекулярных белок-липидных комплексов, причем ВЖК связаны с альбуминами крови, все другие липиды входят в состав лнпопротеидов плазмы крови Любой липопротеид плазмы крови имеют обшие черты строения Наружная оболочка представлена белками, сфинголипидами а так же свободным холистеролом Ядро формируется за счет этерефицированною холистерола и триаци л глицерина Содержание этих компонентов несомненно в разных липопротендах различное. В зависимости от состава липопротеидных частиц они различаются по целому ряду свойств
а) плавучей плотности б) электрофоретическон подвижности, что используется при разделении липопротеидов плазмы крови Липоротеиды ХМ<1гр/л ЛПОНП-1,3-2 гр/л ЛПНП- 3-4,5 гр/л ЛПВП- 2,9-4 гр/л
Содержание липидов в ряду от ХМ до ЛПВП снижается, содержание белков нарастает, постепенно возрастает содержание фосфолипидов возрастает, и только содержание холистерола до лилопротеидов низкой плотности увеличивается, но затем при переходе в лпвп оно снижается. В крови кроме названных частиц могут присутствовать и другие липопротеидьые частицы У здоровых людей всегда обнаруживается фракция липопротеидов промежуточной плотности Эти липопротеидные частицы являются переходными частицами между лпонп и лпнп |Причем это касается их свойств, плотности и тд При патологических состояниях в крови появляется B-лпонп, лпА и др формы липопротеидов Белки содержащиеся в липопротеидах получили название апобелки.
Для лпонп главными - белки апоВ и апоС Для лпвп - белки апоА В то же время в липопротеидных частицах присутствуют в качестве минорных компонентов и другие апобелки Например белки семейства апоЕ присутствуют в лпонп, лпнп Сегодня доказано, что липопротеидны частицы в процессе циркуляции могут обмениваться своими белками. Какую роль выполняют апобелки входящие в состав липопротеидов? Оказывается они выполняют много гранную функцию.
8. Регуляция обмена липидов на уровне организма. Липиды выполняютмножествофункций.Несомненно интенсивность и направленность различных превращений липидов должны соответствовать потребности организма в энергетическом и пластическом материале. Поэтому крайне важны проблемы регуляции.
В ходе мобилизации образуются вжк и глицерол, которые постпают из липоцитов вначале в кровь, а затем в клетки с выделением необходимой клеткам энергии. Этот процесс мобилизации резервных тригяицеридов получил название - липолиз. Он контролируется целым рядом гормонов. К числу их относиться адреналин, норадреналин, глюкагон, соматотропин, кортизол, тироксин, тестостерон. Они стимулируют липолиз.Несомненно есть гормоны стимулирующие процесс липогенеза - инсулин и эстроген. Многие из названных гормонов являются активаторами гормончувствительной липазы лнпоцитов (триацилглицерид-липаза).
Адреналин взаимодействует со своим рецептором на наружной мембране липоцита с образованием гормон-рецептурного комплекса.В ответ на образование гормон-рецепторного комплекса с помощью специального механизма происходит активаци расположенной па внутренней поверхности аденчлатцнклазы. Пока нет гормона аденилатциклаза не активна, не работает. Изменение ее конформации приводит к активации и этот фермент начинает синтезировать из АТФ циклическую АМФ (месенджер). Сам гормон не проникает в клетку.Увеличение внутриклеточной концентрации циклической АМФ активирует фермент протеннкиназу.Этот фермент активирует триацилглщеролмпазу путем ее фосфорилирования т.е. так называемой ковалентной модификацией. Триацилглицероллипаза активна только в фосфорилированной форме. Активация фермента приводит к усилению гидролиза резервных триглицерпдов и к. увеличению выброса влек и глицерола из липоцита в русло крови.
Регуляция пируватдегидрогиназного комплекса, ацетилКоА-карбоксилазы и глицеролфосфатацилтрансферазы осуществляется путем координированного процесса ковалентной модификации, этих ферментов: фосфорилирование - дефосфорилирование.
В целом воздействие инсулина на липоциты приводит во-первых к торможению липолиза в клетках во-вторых активации липогенеза Тем самым способствует накоплению энергетических резервов в организме в виде триацилглиуеринов. Механизм действия эстрогенов и их вляние на липогенез пока еще не изучен окончательно.
9. Паталогия липидного обмена.Нарушение липидного обмена выявляется у людей самыми различными заболеваниями. Эти нарушения можно разделить на первичные и вторичные. При первичных или наследственных нарушениях липидного обмена патологические состояния возникают как следствие генетического дефекта, который сопровождается нарушением синтеза белковых молекул имеющих то или иное отношение к обмену липидов. во-первых это может быть нарушение синтеза белков-рецепторов для ливп во-вторых это может быть нарушение синтеза апопротеинов в-третьнх нарушение синтеза ферментов катализирующих отдельные реакции липидного обмена.
Вторичные нарушения липидного обмена развиваются или в результате имеющегося заболевания, как следствие (сахарный диабет) или как следствие воздействия факторов внешней среды включая сюда и нарушение поведенческих реакций.
Первичные нарушения обмена липидов. К наследственным заболеваниям сопровождающимся нарушением обмена липидов относится
1. гиперхиломикронемия, 2. семейная гиперхолистеринемия 3. болезнь Нимона-Пика 4. болезнь Тея-Сакса
Наследственная гиперхиломикронемия. Болезнь Нимона-Пика В клетках больного отсутствует фермент лизосом - сфингомиелиназа. Или же активность его значительно снижена. Понятно, что в лизосомах накапливается сфингомиелин (т.е.речь идет о типичной болезни накопления). Поражается селезенка, печень, мозг, почки и др. Органы.
Для больных характерно задержка физического и умственного развития, нарушение функций различных органов и в последствии ранняя смерть, обычно в детском возрасте. Болезнь Тея-Сакса. Наследственное нарушение обмена сфиигопипидов. У больных нарушен синтез одного из ферментов расщепляющих ганглиозиды. Особенно много ганглиозидов накапливается в лизосомных клетках головного мозга. Для этих больных так же характерна задержка физического и умственного развития, ранняя смерть. Специфическим признаком этого заболевания является ранняя слепота. Частота врожденных нарушений обмена липидов широко варьирует. Семейная гиперхолистериноемия встречается с частотой 1 на 200 детей. Болезнь Тея-Сакса 1 на 300 родившихся.
Вторичные нарушения обмена липидов. (Желчно-каменная болезнь, атеросклероз) Жировая дистрофия печени (жировая инфильтрация печени)В гепатоцитах накапливаются приемущественно триглицериды, причем масса триглицеридов в тяжелых случаях может составлять до 50% от массы печени (норма не больше 5%).Гепатоциты переполненные трнглицеридами погибают и замещаются фиброзной соединительной тканью, развивается цирроз печени. Ситуация может быть настолько тяжелой, что больные погибают в результате печеночной недостаточности в течении нескольких суток. Это вариант так называемой острой желтой дистрофии печини. Жировая дистрофия печени не является каким-либо специфическим процессом. Оказывается она развивается в ответ на острую или хроническую интоксикацию экзогенного или эндогенного происхождения.
Как известно, расщепление триацилглицеролов происходит под действием фермента липазы.
В полости рта жиры не подвергаются никаким изменениям, т.к. слюна не содержит данный фермент.
Расщепление триацилглицеролов в желудке взрослого человека невелико. Это связано со следующими причинами: во-первых, в желудочном соке содержание липазы крайне низкое; во-вторых, рН желудочного сока (1,5) далек от оптимума действия этого фермента (рН для желудочной липазы 5,5-7,5); в-третьих, в желудке отсутствуют условия для эмульгирования триацилглицеролов, а липаза может активно расщеплять жир, находящийся в форме эмульсии.
Незначительное расщепление триацилглицеролов в желудке облегчает последующее переваривание их в тонком кишечнике под действием липазы панкреатического сока, вырабатываемой поджелудочной железой. Этому процессу способствуют следующие факторы:
– хорошее перемешивание в кишечнике пищевой кашицы с пищеварительными соками. Это связано с тем, что происходит нейтрализация попавшей в кишечник с пищей соляной кислоты желудочного сока бикарбонатами, содержащимися в панкреатическом и кишечном соке. Поэтому выделяющиеся при разложении бикарбонатов пузырьки углекислого газа и способствуют тщательному перемешиванию;
– эмульгирование жира в кишечника. Наиболее мощное эмульгирующее действие на жиры оказывают соли желчных кислот, попадающие в двенадцатиперстную кишку с желчью в виде натриевых солей.
По химической природе желчные кислоты являются производными холановой кислоты и представляют собой конечный продукт метаболизма холестерина. В желчи человека в основном содержатся такие желчные кислоты как: холевая (3,7,12 – триоксихолановая), дезоксихолевая (3,12 – диоксихолановая) и хенодезоксихолевая (3,7 – диоксихолановая).
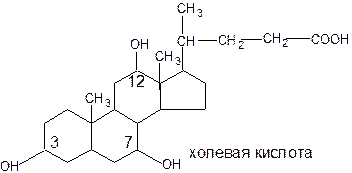
Эти соединения иногда называют парными желчными кислотами, так как они состоят из двух компонентов – желчной кислоты и глицина или таурина:
Считают, что только комбинация соль желчной кислоты + ненасыщенная жирная кислота + моноглицерид придает необходимую степень эмульгирования жира. Соли желчных кислот резко уменьшают поверхностное натяжение на поверхности раздела жир¤вода, благодаря чему они не только облегчают эмульгирование, но и стабилизируют уже образовавшуюся эмульсию;
– активирование панкреатической липазы. Данный фермент является гликопротеином, имеющим мол.массу 48000 и оптимум действия рН 8-9. Как и другие пищеварительные ферменты (пепсин,трипсин и др.) панкреатическая липаза поступает в верхний отдел тонкой кишки в виде неактивной пролипазы. Превращение пролипазы в активную липазу происходит при участии желчных кислот и еще одного белка панкреатического сока – колипазы. Последняя присоединяется к пролипазе в молекулярном соотношении 2:1. Это приводит к тому, что липаза становится активной и устойчивой к действию различных протеолитических ферментов.
Основными продуктами расщепления триацилглицеролов при действии активной панкреатической липазы являютсяβ(2)- моноглицерид и жирные кислоты. Фермент катализирует гидролиз эфирных связей в α(1), α¢(3)-положениях, в результате чего и образуется β(2)-моноглицерол и две молекулы жирной кислоты.
В панкреатическом соке наряду с липазой содержится фермент-изомераза, катализирующая внутримолекулярный перенос ацила из β(2)-положения моноглицерида в α(1)-положение. Поскольку эфирная связь в α-положении чувствительна к действию панкреатической липазы, последняя расщепляет большую часть α-моноглицеридов до конечных продуктов – глицерина и жирной кислоты.
Гидролиз триацилглицеролов при участии панкреатической липазы представлен на рисунке 7.

Рисунок 7–Схема гидролиза триацилглицерола под действием
панкреатической липазы.
Всасывание происходит в проксимальной части тонкой кишки. Основная часть жира всасывается лишь после расщепления его панкреатической липазой на жирные кислоты, моноглицеролы и глицерин. Жирные кислоты с короткой углеродной цепью (менее 10 атомов углерода) и глицерин, будучи хорошо растворимыми в воде, свободно всасываются в кишечнике и поступают в кровь воротной вены, оттуда в печень, минуя какие-либо превращения в кишечной стенке.
Всасывание жирных кислот с длинной цепью и моноглицеролов происходит более сложно. Этот процесс осуществляется при участии желчи и, главным образом, желчных кислот, фосфолипидов, холестерина, входящих в ее состав. Жирные кислоты с длинной цепью и моноглицеролы в просвете кишечника образуют с этими соединениями устойчивые в водной среде мицеллы. Структура мицелл такова, что их гидрофобное ядро (жирные кислоты, моноглицеролы и т.д.) оказывается окруженным снаружи гидрофильной оболочкой из желчных кислот и фосфолипидов.
В составе таких мицелл высшие жирные кислоты переносятся к всасывающей поверхности кишечного эпителия. В результате диффузии, мицеллы проникают в эпителиальные клетки ворсинок, где происходит распад жировых мицелл. При этом желчные кислоты поступают в ток крови и через систему воротной вены попадают сначала в печень, а оттуда вновь в желчь. Таким образом происходит постоянная циркуляция желчных кислот между печенью и кишечником. Этот процесс получил название печеночно-кишечной циркуляции.
После того как произошло всасывание глицерина жирных кислот, они могут использоваться тканями и органами тела в качестве энергетического материала.
Обмен липидов
Липиды являются обязательной составной частью сбалансированного пищевого рациона человека. В среднем в организм взрослого человека с пищей ежесуточно поступает 100 г жиров животного и растительного происхождения. В пожилом возрасте, а также при малой физической нагрузке потребность в жирах снижается, в условиях холодного климата и при тяжелой физической работе увеличивается.
Этапы липидного обмена.
Переваривание и всасывание липидов.
Для переваривания липидов в желудочно-кишечном тракте необходимы, во-первых, липолитические ферменты, во-вторых – эмульгаторы (детергенты), вещества, понижающие поверхностное натяжение и препятствующие склеиванию частиц жира.
Липолитические ферменты – большая группа гидролаз, катализирующих гидролитический распад различных липидов. В желудочно-кишечном тракте находятся липазы – расщепляющие триацилглицерины, фосфолипазы, вызывающие гидролиз фосфолипидов, холестеразы – катализирующие распад эфиров холестерина и др. Максимальную активность названные ферменты проявляют в слабощелочной среде (рН 7,8 – 8,2). Гидролитическому расцеплению подвергаются только эмульгированные жиры.
Примечание: если вещество находится в раздробленном (диспергированном) состоянии и равномерно распределено в массе другого вещества, то такую массу называют дисперсной . Одним из видов дисперсной системы является эмульсия . Она состоит из двух несмешивающихся жидкостей, одна из которых в виде капелек диспергирована в массе другой (капельки жира в молоке). Чтобы получить эмульсию масла в воде, т.е. разделить масло на мельчайшие капельки, необходимо энергично взболтать смесь. Однако при отстаивании капельки масла вновь соберутся вместе над водой, и появится четкая граница по линии раздела двух жидкостей. Для предотвращения склеивания частиц масла добавляют эмульгаторы, которые, окружая каждую капельку масла, препятствуют их склеиванию. В организме основными эмульгаторами являются соли желчных кислот и сывороточный альбумин.
В ротовой полости переваривание жиров не происходит, так как здесь отсутствуют липолитические ферменты. В желудке идет незначительный распад эмульгированного жира пищи (молока, яичного желтка). Это связано с тем, что, во-первых, в желудочном соке хотя и присутствует липаза, но её активность невелика, так как сильнокислая среда желудка (рН 1,0 - 1,5) не соответствует оптимуму рН липазы (7,8 – 8,2), во-вторых, здесь нет эмульгаторов жиров.
Основная масса липидов пищи представлена триацилглицеридами . У взрослого человека основным местом переваривания липидов является тонкий кишечник, где имеются все необходимые для этого процесса условия. Поджелудочная железа и клетки слизистой оболочки кишечника секретируют большую группу липолитических ферментов, а слабощелочная среда обеспечивает их высокую активность. Кроме того, липиды под действием перистальтики кишечника раздробляются (диспергируются) на очень мелкие капли, которые эмульгируются при участии парных желчных кислот и моноацилглицеринов.
В двенадцатиперстной кишке пища подвергается воздействию желчи и сока поджелудочной железы. На первом этапе происходит эмульгирование жира . Эмульсия представляет собой взвесь в водной среде частиц неполярных липидов. По сути дела эмульгирование заключается в дроблении крупных липидных частиц на более мелкие. Происходит этот процесс благодаря трем факторам: 1) перистальтике кишечника, которая способствует перемешиванию и дроблению жировых капель; 2) углекислому газу – он образуется в результате реакции нейтрализации гидрокарбонатов сока кислым содержимым желудка, поступающим туда с пищей; 3) желчным кислотам.
Желчные кислоты образуются в печени из холестерина. Поэтому в основе структуры их молекул лежит циклопентанпергидрофенантрен. В желчи желчного пузыря человека желчные кислоты представлены парными кислотами: гликохолевой, гликодезоксихолевой, гликохенодезоксихолевой, таурохолевой, тауродезоксихолевой и таурохенодезоксихолевой кислотой. Парные желчные кислоты представляют собой соединения кислот с глицином и таурином и образуются в желчном пузыре. Образующиеся желчные кислоты поступают из печени в двенадцатиперстную кишку с желчью. В нейтральной или слабощелочной среде просвета кишечника желчные кислоты, в основном таурохолевая и гликохолевая, являются амфифильными и эмульгирующими агентами, а также стабилизаторами образующейся эмульсии.
Роль желчных кислот и солей:
- Эмульгирование липидов.
- Активация липолитических ферментов.
- Образование простой мицеллы.
- Образование смешанной мицеллы.
- Всасывание липидов и жирорастворимых витаминов в лимфатическую систему.
- Выведение из организма холестерина.
Взаимодействуя гидрофобными частями своих молекул с жиром, а гидрофильной, полярной частью – с водным содержимым кишечника, желчные кислоты способствуют дроблению жира на мелкие частицы, т.е. эмульгированию. Стабилизирующее влияние желчных кислот на эмульсионные частицы обусловлено тем, что они препятствуют конгломерации (слипанию) эмульсионных частиц. Желчные кислоты покрывают поверхность эмульсионной частицы в виде монослоя. При этом наружу, к водному содержимому, направлены полярные части молекул желчных кислот. В результате поверхность частицы приобретает суммарный электрический заряд, который будет одноименным у всех других эмульсионных частиц. В силу электростатического взаимодействия между отдельными частицами возникает отталкивание.
Так как липиды в основном нерастворимы в воде, то они подвергаются действию гидролитических ферментов только на границе раздела между липидами и водной фазой. Скорость реакции, помимо других факторов, зависит от площади этой границы раздела. Поэтому чем выше степень эмульгирования и чем меньше отдельные липидные капли. Тем больше величина общей доступной поверхности.
В соке поджелудочной железы присутствует предшественник липазы , активирующийся в просвете кишечника. Активный фермент ускоряет реакцию гидролиза ацилглицеридов. Гидролиз ТАГ сначала происходит в положении 1 или 3, что приводит к образованию диацилглицеридов, которые затем гидролизуются до 2-моноацилглицеридов. Меньшая часть (40%) моноацилглицеридов подвергается гидролизу до глицерина. Для остальной части процесс ферментативного гидролиза завершается на этапе образования 2-моноацилглицеридов.
В соке поджелудочной железы присутствуют и другие ферменты, способные расщеплять липиды. В частности, эстеразы катализируют преимущественно гидролиз эфиров жирных кислот. В поджелудочной железе синтезируется профосфолипаза А 2 . Фермент катализирует отщепление молекулы жирной кислоты от фосфатидилхолина с образованием лизофосфатидилхолина.
Переваривание фосфолипидов обеспечивается группой фосфолипаз, которые последовательно гидролизуют молекулу на составные части. Вначале фосфолипаза А отщепляет кислоту от второго углеродного атома глицерина. Оставшаяся часть молекулы называется лизофосфолипидом и обладает свойствами эмульгатора. Затем последовательно действуют фосфолипазы В, С, D, которые расщепляют молекулу на глицерин, жирную кислоту, фосфорную кислоту и азотистое основание. Холестерин, находящийся в пище в виде эфиров, распадается под действием холестераз.
Оставшееся количество непереваренных липидов либо всасывается в тонкой кишке, либо поступают в толстую кишку и выводятся с калом.
Процесс всасывания характеризуется тем, что водорастворимые продукты распада (глицерин, моноацилглицерины, фосфорная кислота, азотистые основания) легко проникают в клетки слизистой оболочки кишечника. Жирорастворимые компоненты (жирные кислоты, холестерин и др.) всасываются при участии парных желчных кислот, с которыми они образуют водорастворимые комплексы.
Продукты ферментативного гидролиза жира взаимодействуют с водной средой, объединяются в мельчайшие частицы – мицеллы . По размеру они гораздо меньше, чем эмульсионные. Снаружи мицеллы, подобно эмульсионным частицам, покрыты слоем желчных кислот. Т.е. в просвете кишечника парные желчные кислоты соединяются в мицеллы (очень маленькие капельки), наружная часть которых образована гидрофильными частями этих кислот, а внутренняя – гидрофобными. Жирные кислоты, холестерин и другие жирорастворимые вещества проникают во внутреннюю часть мицеллы и в ее составе всасываются в клетки слизистой оболочки кишечника, где мицеллы распадаются.
Основная часть мицелл целиком или после предварительного (пристеночного) разрушения всасывается через стенку тонкого кишечника. Желчные кислоты проходят через барьер слизистой оболочки в связанном с липидами состоянии. В дальнейшем по венам кишечника они поступают, а портальный кровоток, оттуда извлекаются печенью и затем снова поступают с желчью в двенадцатиперстную кишку. В результате такого кругооборота небольшое количество желчных кислот обеспечивает всасывание достаточно больших количеств жиров.
Всосавшиеся продукты расщепления липидов в клетках слизистой оболочки кишечника подвергаются процессам ресинтеза . Там имеются ферментные системы, которые могут превращаться свободные жирные кислоты, моно- и диацилглицериды в триацилглицериды.
Таким образом, в результате процессов всасывания в клетках слизистой оболочки тонкой кишки накапливаются все конечные продукты распада липидов. Здесь же в стенке кишки из этих продуктов происходит первичный синтез липидов, но уже в специфичных для данного организма (триацилглицеридов, фосфолипидов, эфиров холестерина и др.).











